
Protistoj
| Protistoj | ||||||
|---|---|---|---|---|---|---|

| ||||||
Sunbestetoj (Actinophrys sol) | ||||||
| Biologia klasado | ||||||
| ||||||
| Subregno | ||||||
|


Protistoj estas eŭkariotaj organismoj, kiuj hodiaŭ estas kalkulitaj kiel unu regno de la organismoj kaj ne al la plantoj, bestoj aŭ fungoj. Ilia plimulto estas unuĉelulo, kvankam kelkajn pli grandan multĉelulan organismon oni kalkulas al protistoj.
Al la protistoj apartenas algoj, mukfungoj kaj unuĉelulaj protozooj.
Priskribo
[redakti | redakti fonton]La protistoj estas interne – kiel ĉiu organismo – per citoplasmo (kiel viva materialo) plenigita kaj ekstere per ĉelmembrano ĉirkaŭvolvita. En la citoplasmo troviĝas plej ofte ĉelkerno por la direkto de la vivfenomenoj; foje troviĝas tie pliaj ĉelkernoj. Ĉe iuj protistoj formiĝas ekstere ankoraŭ ŝelo-estaĵoj; multaj kapablas formi cistovualon? (aŭ cistoŝelon), por travivi la malkonvenajn mediajn kondiĉojn. La ĉeloj havas plej ofte tre etan grandon, kaj nur kelkaj atingas la diametron de kelkaj milimetroj aŭ eĉ centimetroj.
La antaŭen-moviĝo okazas ofto naĝante per flageloj aŭ cilioj aŭ per formo de ŝajnpiedoj. Kelkaj specioj ŝvebas en la akvo simple nur per longaj ĉelaj eletendiĝoj.
La plimulto de la protistoj vivas en maro, parte proksime al la surfaco, parte ŝvebante en la akvo, parte surfunde rampante, parte sur ŝtonoj, plantoj k.t.p. fikse sidante; aliaj troviĝas en dolĉakvo, malmultaj surtere. Tiel formas la tribo de verdalgoj plimulton de en la dolĉakvo vivantaj fitoplanktonoj. La tribo de sulkoalgoj (kirasalgoj) donas la ĉefmason de la maraj fitoplanktonoj. Ekzistas heterotrofaj, aŭtotrofaj, aerobiaj kaj anaerobiaj formoj. Kelkaj protistoj vivas ankaŭ parazite en la pli evoluintaj bestoj. Multaj aperas en mirakla grandnombro, kaj la neputriĝantaj postrestaĵoj de la formortitaj reprezentantoj – kiel la silikaj ŝeloj de la radiobestetoj aŭ la kalkaj ŝeloj de ĉambretaj rizopodoj (truŝelaj, foraminiferoj) (Foraminifera) – formas kelkloke tutan montaran tavolon.
La protistoj multobliĝas per la kutima neseksa duoniĝo (vidu mitozo). Ĉe kelkaj specioj ekzistas ankaŭ multobliĝoj, ĉe aliaj seksaj procesoj (vidu izogamio, konjugacio).
Dum la evoluado formiĝis el la praaj protistoj ĉiu pli evoluinta plurĉela organismo, verŝajne tra la vojo de formado de ĉelkolonioj, kio estas ankoraŭ hodiaŭ spektabla ĉe kelkaj algoj.
Ĉar la plej gravaj vivfenomenoj de la protistoj – kiel moviĝo, ekscitebleco, pliiĝo - estas tre bone studeblaj, ili estas ŝatataj objektoj de la biologia esplorado.
Trajtoj
[redakti | redakti fonton]Ĉar Protistoj ne estas grupo monofiletika, maleblas specifi trajtojn kiuj povas difini aŭ distingi are tiujn vivajn estaĵojn. La komunaj trajtoj estas propraj de la eŭkariotoj ĝenerale, kaj tial ili estas kunhavataj, scepte pro duaranga perdo, fare de plantoj, animaloj aŭ fungoj. La suba listo montras ĉefe enorman diversecon:
- Habitato: Neniu el la anoj de la grupo estas tute adaptita al la ekzistado en aero, tial ke tiuj kiuj ne estas rekte akvoloĝantoj, disvolviĝas ĉu en surteraj medioj malsekaj ĉu en la interna medio de aliaj organismoj.

- Ĉelorganizado: Eŭkariotoj (ĉeloj kun nukleo), unuĉeluloj aŭ plurĉeluloj. La plej grandaj, nome la brunalgoj de la genro Laminaria, povas esti dekojn da metroj longaj, sed inter la cetero hegemonias la mikroskopaj formoj.
- Strukturo: Kutime oni asertas, ke ne ekzistas histoj en ajna protisto, sed ĉe la brunalgoj (feoficeaj) la komplekseco atingas nivelon tre proksiman al hista nivelo, inklude la ekzistado de plasmodesmoj (p. ekz., ĉe la brunalgo Egregia). Multaj de la protistoj kiuj estas plurĉelulaj disponas de ĉelmuroj de varia kompono, kaj la unuĉeluloj memtrofaj ofte estas ŝirmitaj per "teko", kiel okazas ĉe la rimarkinda kazo de la diatomeoj, aŭ tiuj kiuj disponas de skvamoj aŭ plifortigiloj. La predantaj unuĉeluloj (fagotrofoj) kutime montras nudajn ĉelojn (sen ŝirmigiloj). La unuĉelaj formoj ofte disponas de moveblecon pere de rampado aŭ, pli ofte, per apendicoj de diversaj tipoj nomitaj cilioj kaj flageloj.
- Manĝomaniero: Protistoj povas esti memtrofuloj (per fotosintezo), heterotrofuloj aŭ miksotrofuloj. La heterotrofuloj povas esti tio per ingesto (fagotrofuloj) aŭ per osmoza absorbado (osmotrofuloj). Kelkaj estas parazitaj, kiel ekzemple la apikompleksuloj kaj la Trypanosoma (tripanosomoj), kiuj okazigas tre aĉajn malsanojn al homaj estaĵoj.
- Metabolo de oksigeno: Kiel propras ĉe la eŭkariotoj, la protistoj estas aerobiaj (tio estas, ili uzas oksigenon por ekhavi la energion de la organikaj substancoj), sed kelkaj estas duarange anaerobiaj, estiĝinte adaptitaj al medioj malabundaj je tiu substanco.
- Reproduktado kaj disvolviĝo: Ĉe protistoj eblas neseksa (klona) kaj seksa reproduktadoj, per gametoj, ofte alternante neseksan kaj seksan en la sama specio. La plurĉelaj algoj ekzemple montras ofte "alternado de generacioj". Ĉiuokaze ne ekzistas ajna embrio.
- Ekologio: La protistoj estas inter la plej gravaj komponantoj de la planktono (organismoj kiuj vivas flosante en akvo), de la bentoso (de la fundo de akvaj ekosistemoj) kaj de la edafo (de la komunumo kiu vivas en la grundoj). Estas multaj gravaj tipoj kaj de parazitismo kaj de mutualismo. Tiuokaze menciendas la Mastigophora (flageluloj), kiuj intervenas en la digesto de la ligno fare de la termitoj, aŭ tiuj kiuj vivas en la rumeno de la bovoj. La alga simbionto de la likenoj estas preskaŭ ĉiam verda algo unuĉela.
Klasigo
[redakti | redakti fonton]La regno Protista ludis gravan rolon en la origino kaj evoluo de la eŭkariota ĉelo. Ĝi konstituas taksonon parafiletikan rilate al aliaj regnoj, ĉar ĝi baziĝas sur la pleziomorfa karaktero de la unuĉeleco kaj ne enhavas ĉiujn posteulojn de la specioj kiujn ĝi enhavas. Eĉ tial, estas inter ili membroj kiuj pliigis sian nivelon de komplekseco direkte al la plurĉeleco. Kelkaj grupoj de eŭkariotoj disvolvis sendepende la plurĉelecon deveninte el protisto: animaloj, fungoj, plantoj, musko-ŝimoj kaj brunalgoj. El ili, la tri unue menciitaj grupoj estas konsiderataj sendependaj regnoj, dum la du laste menciitaj grupoj estas inkluditaj ene de la regno Protista.
La klasigo de la regno Protista multe antaŭeniris en la lastaj jardekoj kaj pro la ultrastrukturaj studoj kaj pro la genetikaj analizoj. La morfologiaj kaj funkciaj trajtoj havigas ideon pri la diverseco de la protistoj kaj pri ĝia evoluego, tio estas, la ŝanĝoj kiuj tuŝas ilian korpoplanon kaj iliajn manierojn survivi kaj reproduktiĝi. Por ekzemplo, la interna skeleto aŭ ĉelskeleto havigas lumon por la ideo de la baza korporplano de la organismo, ĉar ĝi funkcias kiel eltenilo kaj devas esti sufiĉe plastija por modifiĝi cele al la movebleco kaj al la manĝado. Uzante tiun alproksimigon, Cavalier-Smith[1][2][3][4][5][6] establis malgrandan nombron de filumoj kiuj, kvankam ili ne estas enhavemaj kaj diversaj, ja ili havas unuigan trajton en la baza korporplano, simile al ti kio ojazas ĉe la tradiciaj filumoj de aliaj regnoj, kiel ĉe ĥorduloj, moluskoj, artropodoj (en animaloj), aŭ traĥeofituloj (en plantoj). Kelkaj el tiuj analizoj utilis por dedukti kia estus estinta la pra-eŭkarioto: nome unuĉela, flagelhava kaj fagotrofulo (trajto per kiu ĝi povi engluti la pra-mitokondrion, kio ebligis la spiradon aerobian kaj poste la pra-kloroplaston, kio rezultis en la apero de organismoj eŭkariotaj fotosintezaj).
Ĉefaj tipoj
[redakti | redakti fonton]Protistoj montras larĝan gamon de apartaj morfologiaj tipoj kiuj estis uzitaj por klasifiki ilin por praktikaj celoj, kvankam la plej multaj el tiuj kategorioj ne reprezentas evoluajn koheziajn genliniojn aŭ kladojn kaj anstataŭe evoluis sendepende plurajn fojojn. La plej rekoneblaj tipoj estas jenaj:[7]
- Amoeba. Karakterizitaj pro siaj neregulaj, flekseblaj formoj, tiuj protistoj moviĝas per etendado de partoj de sia citoplasmo, konataj kiel ŝajnpiedoj aŭ pseŭdopodoj, por rampi sur surfacoj.[8] Multaj grupoj de ameboj estas nudaj, sed esploritaj ameboj kaj foraminiferoj kreskas ŝelon ĉirkaŭ sia ĉelo faritan el digestita materialo aŭ ĉirkaŭaj derompaĵoj. Kelkaj, konataj kiel radiuloj kaj heliozooj, havas specialajn sferajn formojn kun mikrotubul-subtenataj pseŭdopodoj radiantaj de la ĉelo.[7] Kelkaj ameboj estas kapablaj je produktado de persekutitaj multĉelaj stadioj kiuj portas sporojn, ofte per agregado kune; tiuj estas konataj kiel ŝlimo-ŝimoj.[9] La ĉefaj kladoj enhavantaj amebojn estas Amoebozoa (inkluzive de diversaj ŝlimoŝimoj kaj testataj ameboj) kaj Rhizaria (inkluzive de famaj grupoj kiel ekzemple foraminiferoj kaj radiuloj, same kiel kelkaj esploritaj ameboj).[10][11] Eĉ iuj individuaj ameboj povas kreski ĝis gigantaj grandecoj videblaj al la nuda okulo.[12][13]
- Flagellata. Tiuj protistoj estas provizitaj per unu aŭ pluraj vip-similaj alpendaĵoj nomitaj cilioj, undulipodioj aŭ eŭkariotaj flageloj, kiuj ebligas ilin naĝi aŭ gliti libere tra la medio. La esprimoj "cilio" kaj "eŭkariota flagelo" estas interŝanĝeblaj de biologia perspektivo. Tamen ilia uzado dependas de la aŭtoro: iuj preferas rezervi ciliojn por pli mallongaj alpendaĵoj kaj flagelojn por pli longaj, dum aliaj preferas ciliojn por eŭkariotoj kaj flagelojn por prokariotoj. La esprimo "undulipodio" estis proponita por unuigi la du konceptojn, ĉar ĝi rilatas specife al la homologa mikrotubeta strukturo trovita en ambaŭ, sed ne trovita en prokariotaj flageloj.[14][15][16] Flagelatoj troviĝas en ĉiuj genlinioj, reflektante ke la komuna prapatro de ĉiuj vivantaj eŭkariotoj estis flagelato. Ili kutime elmontras du ciliojn (ekz., en Provora, Telonemia, Stramenopiles, Alveolata, Obazoa kaj la plej multaj Elfosuloj), sed ekzistas kelkaj flagelaj grupoj kun alta nombro da cilioj (kiel ekzemple Hemimastigophora kaj aliaj Elfosuloj).[7] Kelkaj grupoj, kiel la konataj Ciliuloj kaj la parazitaj Opalinidae, havas ĉelan surfacon kovritan per vicoj de cilioj, kiuj batas ritme. Kelkaj grupoj de ameboj konservis siajn flagelojn, igante ilin Amoeboflagellate.[17]
- Algoj. Ili estas la fotosintezaj protistoj, kaj povas troviĝi en la plej multaj el la ĉefaj kladoj, tute intermiksitaj kun heterotrofaj protistoj kiuj estas tradicie nomitaj protozooj.[18] Algoj elmontras la plej diversan gamon da morfologioj, de unuopaj flagelitaj aŭ kokoidaj ĉeloj (ekz., kriptofitoj, haptofitoj, dinoflagelatoj, kromeridoj, multaj verdaj algoj, okrofitoj, eŭglenofitoj) ĝis ameboidaj ĉeloj (kloraĥniofitoj) ĝis koloniaj kaj multĉelaj makroskopaj formoj (ruĝaj algoj, kelkaj verdaj algoj, kaj kelkaj okrofitoj kiel ekzemple Laminariales).[19]
- Fungo-similaj protistoj. Pluraj kladoj de protistoj evoluigis aspekton similan al fungoj tra hif-similaj strukturoj kaj saprofita nutrado. Ili evoluis plurfoje, ofte tre for de veraj fungoj (ekz., la oomicetoj, labirintulomicetoj kaj hifokitridoj, en Stramenopiles; la miksomicetoj, en Amoebozoa; la fitomiketoj, en Rhizaria; la perkinsozooj, en Alveolata).[20][21]
- Sporozoa. Ĉi tiu kategorio tradicie inkludis parazitajn protistojn kiuj reproduktiĝis per sporoj (nome la apikompleksoj, mikrosporidoj, miksozooj kaj ascetosporeoj).[22] Ĝia nuna uzo estas limigita al la apikompleksoj,[23] kiel Plasmodium falciparum, la kaŭzo de malario.[24]
Taksonoj
[redakti | redakti fonton]- Stramenopiles
- Alveolata
- Rhizaria
- Excavata
- Amoebozoa
- Opisthokonta
Ekzemploj
[redakti | redakti fonton]En la jenaj fotoj oni montras reprezentanton de ĉiu el ĉefaj filumoj de protistoj.
-
 Macrocystis pyrifera (Heterokonta)
Macrocystis pyrifera (Heterokonta) -
 Paramecium aurelia (Ciliophora)
Paramecium aurelia (Ciliophora) -
 Plasmodium (Apicomplexa)
Plasmodium (Apicomplexa) -
 Dinophysis (Dinoflagellata)
Dinophysis (Dinoflagellata) -
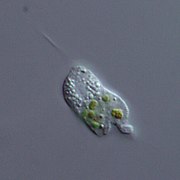 Cercomonas (Cercozoa)
Cercomonas (Cercozoa) -
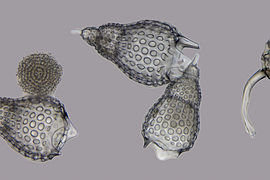 Radiolario (Radiolaria)
Radiolario (Radiolaria) -
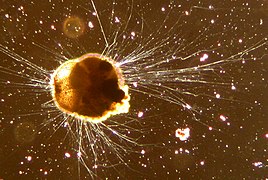 Ammonia tepida (Foraminifera)
Ammonia tepida (Foraminifera) -
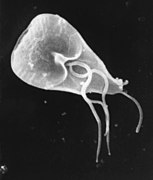 Giardia lamblia (Metamonada)
Giardia lamblia (Metamonada) -
 Trachelomonas (Euglenozoa)
Trachelomonas (Euglenozoa) -
 Percolomonas (Percolozoa)
Percolomonas (Percolozoa) -
 Amoeba proteus (Amoebozoa)
Amoeba proteus (Amoebozoa) -
 Nuclearia (Choanozoa)
Nuclearia (Choanozoa) -
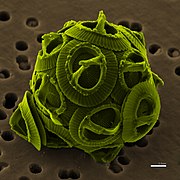 Gephyrocapsa oceanica (Haptophyta)
Gephyrocapsa oceanica (Haptophyta) -
 Rhodomonas salina (Cryptophyta)
Rhodomonas salina (Cryptophyta) -
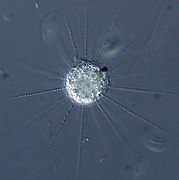 Raphidiophrys contractilis (Centrohelida)
Raphidiophrys contractilis (Centrohelida)














Biologio
[redakti | redakti fonton]Ĝenerale, protistoj havas tipajn eŭkariotajn ĉelojn kiuj sekvas la samajn principojn de biologio priskribitajn por tiuj ĉeloj ene de la "pli altaj" eŭkariotoj (bestoj, fungoj kaj plantoj).[25] Tamen, multaj evoluigis diversajn unikajn fiziologiajn adaptiĝojn kiuj ne aperas en la ceteraj eŭkariotoj,[26] kaj fakte protistoj ampleksas preskaŭ la tutan larĝan spektron de biologiaj karakterizaĵoj atenditaj ĉe eŭkariotoj.[27]
Nutrado
[redakti | redakti fonton]Protistoj montras vastan gamon de manĝpreferoj kaj manĝmekanismojn.[28][29] Laŭ la fonto de iliaj nutraĵoj, ili povas esti dividitaj en aŭtotrofoj (produktantoj) kaj heterotrofoj (konsumantoj). Aŭtotrofaj protistoj sintezas siajn proprajn organikajn substancojn el neorganikaj substratoj tra la procezo de fotosintezo, uzante lumon kiel la fonton de energio;[30] sekve, ili ankaŭ estas konataj kiel fototrofoj.[31]
Heterotrofaj protistoj akiras organikajn molekulojn sintezitaj de aliaj organismoj, kaj povas esti plu dividitaj laŭ la grandeco de siaj nutraĵoj. Tiuj kiuj manĝas solveblajn molekulojn[30] aŭ makromolekulojn malpli ol 0.5 μm en grandeco estas nomitaj osmotrofoj,[29] kaj ili absorbas ilin per difuzo, ciliarfosaĵoj, transportproteinoj de la ĉelmembrano, kaj speco de endocitozo (t.e., trudado de la ĉelmembrano en vakuolojn, nomitajn endozomoj) konata kiel pinocitozo[28] aŭ fluid-faza endocitozo.[29] Tiuj kiuj manĝas organikajn partiklojn pli ol 0,5 μm en grandeco aŭ tutajn ĉelojn estas nomitaj fagotrofoj, kaj ili konsumas ilin tra speco de endocitozo konata kiel fagocitozo.[29][30] Endocitozo estas konsiderata unu el la plej gravaj adaptiĝoj en la origino de eŭkariotoj, ĉar ĝi pliigis la eblan manĝprovizon, kaj fagocitozo permesis la endosimbiozon kaj evoluon de mitokondrioj kaj kloroplastoj. En kaj osmotrofoj kaj fagotrofoj, endocitozo ofte estas limigita al specifa regiono de la ĉelmembrano, konata kiel la citostomo, kiu povas esti sekvita per citofaringo, specialeca vojo apogita per mikrotubetoj.[29]
Osmotrofio
[redakti | redakti fonton]Osmotrofaj protistoj akiras solveblajn nutraĵojn per membran-kanaloj kaj transportiloj, sed ankaŭ per malsamaj specoj de pinocitozo. Makropinocitozo implikas la faldadon de membrano en rufojn,[32] kio kreas grandajn (0,2 ĝis 1,0 μm) vakuolojn. Mikropinocitozo implikas pli malgrandajn vezikojn kiuj estas kutime formitaj per klatrino. En ambaŭ kazoj, la vezikoj kunfandiĝas en digesta vakuolo aŭ endozomo kie okazas digestado.[29] Kelkaj osmotrofoj, nomitaj saprotrofoj aŭ lizotrofoj, elfaras eksteran digestadon liberigante enzimojn en la medion kaj putrigante organikan materion[28] en pli simplajn molekulojn kiuj povas esti absorbitaj. Tiu ekstera digestado havas klaran avantaĝon: ĝi ebligas pli grandan kontrolon de la substancoj kiuj povas eniri en la ĉelon, tiel minimumigante la konsumadon de damaĝaj substancoj aŭ infekton.[33]
Probable ĉiuj eŭkariotoj estas kapablaj je osmotrofio, sed kelkaj ne havas alternativon akiri nutraĵojn. Devigaj osmotrofoj kaj saprotrofoj inkluzivas kelkajn eŭglenidojn, kelkajn verdajn algojn, la homan paraziton Blastocystis, kelkajn metamonadojn,[28] la parazitajn tripanosomatidojn,[34] kaj la fungosimilajn oomicetojn kaj hifokitridojn.[33]

Fagotrofio
[redakti | redakti fonton]Fagotrofia nutrado konsistas el du fazoj: la koncentriĝo de manĝpartikloj en la medio, kaj la fagocitozo, kiu enfermas la manĝpartiklon en vakuolo (la fagozomo)[29] kie okazas digestado. Ĉe ciliuloj kaj plej fagotrofaj flagelatoj, digestado okazas ĉe la buŝa regiono aŭ citostomo, kiu estas kovrita per ununura membrano el kiu formiĝas vakuoloj; la fagosomoj tiam povas esti transportitaj al la interno de la ĉelo laŭ la citofaringo.[36] Ĉe ameboj, fagocitozo okazas ie ajn sur la ĉela surfaco. La meza manĝpartikla grandeco estas ĉirkaŭ unu dekono de la grandeco de la protista ĉelo.[37]
Fagotrofaj protistoj povas esti plue klasifikitaj laŭ kiel ili alproksimiĝas al la nutraĵoj. La filtromanĝantoj akiras malgrandajn, suspenditajn manĝpartiklojn aŭ prokariotajn ĉelojn kaj akumulas ilin per filtrado en la citostomon (ekz., koanoflagelatoj, kelkaj krizomonadoj (flavaj algoj), la plej multaj ciliuloj);[28] filtromanĝaj flageluloj akumulas partiklojn propulsante ilin per flagelo tra kolumo de rigidaj tentakloj aŭ pseŭdopodoj kiuj funkcias kiel filtrilo, dum filtromanĝantaj ciliuloj generas akvofluojn pere de cilioj kaj membranetzonoj ĉirkaŭantaj la citostomon. La rabomanĝantoj (ekz., bikozoecidoj, krizomonadoj, kinetoplastidoj, kelkaj eŭglenidoj, multaj dinoflagelatoj kaj ciliuloj), anstataŭ reteni ĉiujn partiklojn en groco, kaptas ĉiun partiklon unuope.[37]

Inter rabantaj protistoj, la paŝtistoj serĉas kaj konsumas predon el surfacoj kovritaj per eblaj nutraĵoj kiel bakteriaj gazonoj, dum la predantoj aktive persekutas malabundan predon.[28] Predantoj kiuj manĝas filamentajn algojn aŭ fungajn hifojn ĉu glutas la filamentojn tute aŭ penetras la ĉelan muron kaj ingestas la citoplasmon (ekz., Viridiraptoridae).[28] Predantoj povas havi adaptiĝojn por ĉasi predon, kiel ekzemple "toksistoj" kiuj senmovigas predĉelojn. Certaj ciliuloj evoluigis specializitan specon de rapta manĝado nomita histofagio, laŭ kiu ili atakas difektitajn sed vivantajn bestojn (ekz., anelidoj kaj malgrandaj krustacoj), eniras la vundojn, kaj ingestas bestan histon. Grandaj raptoraj ameboj enfermas sian predon en "manĝotaso" de pseŭdopodoj, antaŭ la formado de la manĝvakuolo.[37] Finfine, disvastigmanĝantoj (ekz., heliozooj, foraminiferoj kaj multaj aliaj ameboj, suktoriaj ciliuloj) englutas predojn kiuj hazarde kolizias kun siaj pseŭdopodoj aŭ, en la kazo de ciliuloj, ili uzas tentaklojn kiuj portas toksistojn aŭ ekstruzomojn por senmovigi la predon.[37]
Konsumantoj de prokariotoj estas populare nomitaj bakterivoruloj (ekz., la plej multaj ameboj), dum konsumantoj (inkluzive de osmotrofaj parazitoj) de eŭkariotoj estas konataj kiel eŭkariovoruloj. Aparte, eŭkariovoruloj kiuj manĝas unuĉelaj protistoj estas citotrofoj (ekz., kolponemidoj, kolpodelidoj, multaj ameboj, kelkaj ciliuloj); tiuj kiuj manĝas fungojn estas mikofagoj aŭ mikotrofoj (ekz., la ciliulfamilio Grossglockneriidae de premitaj mikofagoj);[38] tiuj kiuj kaptas nematodojn estas nematofagoj;[39] kaj tiuj kiuj manĝas algojn estas fikotrofoj (ekz., vampirelidoj).[28]
Miksotrofio
[redakti | redakti fonton]Plimulto de aŭtotrofaj protistoj estas miksotrofoj[40] kaj kombinas fotosintezon kun fagocitozon. La terminoj "miksotrofo" kaj "miksoplanktono" preskaŭ nur referencas al protistoj kiuj plenumas fotosintezon kaj fagocitozon (foto-fagotrofoj). Osmotrofio ĉiam estas, sed ne estis atentita. Kiel tio, "puraj" fototrofoj (nekapablaj je fagocitozo) kaj "puraj" fagotrofoj (nekapablaj je fotosintezo) estas teknike miksotrofaj pro sia esenca kapablo por osmotrofio, sed kutime ne oni informas tiukadre.[41] Ili estas klasitaj en variaj funkciaj grupoj aŭ 'miksotipoj'.[42][43] Konstituaj miksotrofoj havas esencan kapablon por fotosintezo pere de jam ĉeestaj kloroplastoj, kaj havas diversajn manĝokutimojn, ĉar kelkaj postulas fototrofion, aliaj fagotrofion, kaj aliaj estas premitaj miksotrofoj (ekz., nanoflageluloj same kiel kelkaj haptofitoj kaj dinoflageluloj). Nekonstituaj miksotrofoj akiras la kapablon fotosintezigi elprenante kloroplastojn el sia predo, laŭ procezo konat kiel kleptoplastio. Nekonstituaj povas esti dividitaj en du grupoj, nome la jenaj: ĝeneralistoj, kiuj povas elpreni kloroplastojn el vario de predoj (ekz., oligotriĥaj ciliuloj), aŭ specialistoj, kiuj povas nur akiri kloroplastojn el kelkaj malmultaj specifaj predoj (ekz., Rapaza viridis povas elpreni nur el ĉeloj de Tetraselmis).[44] La specialistoj sievice estas dividataj en du tipoj: plastidaj, kiuj enhavas diferencajn plastidojn (ekz., Mesodinium, Dinophysis), kaj endosimbiozaj, kiuj enhavas ĉiujn endosimbiotojn (ekz., miksotrofa Rhizaria same kiel Foraminiferoj kaj Radiuloj, dinoflageluloj kiel Noctiluca).[42]
Inter ekskluzive heterotrofaj protistoj, oni observis ankaŭ varion de manĝoreĝimoj. La diplonemidoj, kiuj enloĝas profundajn akvojn kie fotosintezo forestas, povas flekseble ŝanĝi inter osmotrofio kaj bakterivoreco depende de la mediaj kondiĉoj.[45]
Osmoregulado
[redakti | redakti fonton]
Multaj nesalakvaj protistoj bezonas osmoreguligi (t.e., forigi troan akvovolumon por alĝustigi la jonajn koncentrojn) ĉar nesala akvo eniras troe per osmozo el la medio[46] kaj per endocitozo se temas pri manĝado.[36] Osmoreguligo estas farata pere de aktivaj jontransportiloj de la ĉelmembrano kaj pere de kuntiriĝemaj vakuoloj, nome specializitaj organetoj kiuj periode ekskrecias fluaĵon riĉa je natrio kaj kalio pere de ciklo de diastolo kaj sistolo. La ciklo haltiĝas kiam la ĉeloj estas metitaj en medio kun diferenca salenhavo, ĝis la ĉelo adaptiĝas.[26]
La kuntiriĝemaj vakuoloj estas ĉirkaŭitaj de la spongiomo, aro de citoplasmaj vezikoj aŭ tuboj kiuj malrapide kolektas likvaĵon de la citoplasmo en la vakuolo. La vakuoloj tiam kuntiriĝas kaj eligas la likvaĵon ekstere de la ĉelo tra poro. La kuntiriĝa mekanismo varias depende de la protisto: en ciliuloj, la spongiomo estas kunmetita de neregulaj tubetoj kaj aktinaj filamentoj serpentumas ĉirkaŭ la poro kaj super la vakuolsurfaco, kune kun mikrotubetoj; en la plej multaj flageluloj kaj ameboj, la spongiomo estas kunmetita de kaj vezikoj kaj tubuloj; en dinoflageluloj, flagela radiketo disbranĉiĝas por formi kuntiriĝan ingon ĉirkaŭ la vakuolo (konata kiel pusulo).[36] Ankaŭ la loko kaj kvanto varias: unuĉelaj flagelaj algoj (kriptomonadoj, eŭglenidoj, prazinofitoj, flavaj algoj, haptofitoj, ktp.) havas tipe ununuran kuntiriĝeman vakuolon en fiksa pozicio; nudaj ameboj havas multajn malgrandajn vezikojn kiuj kunfandiĝas en unu vakuolo kaj poste fendetiĝas denove post sekrecio. Maraj aŭ parazitaj protistoj (ekz., metamonadoj), same kiel tiuj kun rigidaj ĉelaj muroj, ne havas tiujn vakuolojn.[46]
Spirado
[redakti | redakti fonton]La lasta eŭkariota komuna praulo estis aerobia, portanta mitokondriojn por oksida metabolo. Multaj genlinioj de libervivaj kaj parazitaj protistoj sendepende evoluis kaj adaptiĝis por enloĝi neaerobiajn aŭ mikroaerofilajn vivejojn, per modifado de la fruaj mitokondrioj en hidrogenosomojn, organetojn kiuj generas ATP neaerobie tra fermentado de piruvato. Paralele, ĉe la mikroaerofilaj tripanosomatidaj protistoj, la fermenta glikozomo evoluis el la peroksisomo.[26]
Sensa percepto
[redakti | redakti fonton]
Multaj flageluloj kaj verŝajne ĉiuj movaj algoj elmontras pozitivan fototakson (t.e. ili naĝas aŭ glitas direkte al fonto de lumo).
Por tiu celo, ili elmontras tri specojn de fotoreceptoroj aŭ "okuliloj": (1) receptoroj kun malpezaj antenoj, troveblaj en multaj verdaj algoj, dinoflageluloj kaj kriptofitoj; (2) riceviloj kun netravideblaj ekranoj; kaj (3) kompleksaj ocelloidoj kun intraĉelaj lensoj, troveblaj en unu grupo de predantaj dinoflageluloj, nome la Warnowiaceae.
Aldone, kelkaj ciliuloj orientiĝas rilate al la gravita kampo de la Tero dum moviĝado (geotaxis), kaj aliaj naĝas rilate al la koncentriĝo de dissolvita oksigeno en la akvo.[26]
Endosimbiontoj
[redakti | redakti fonton]Protistoj havas markitan tendencon inkludi endosimbiotojn en siaj ĉeloj, kaj tiuj produktas novajn fiziologiajn ŝancojn.
Kelkaj asocioj estas pli konstantaj, kiel Paramecium bursaria kaj ĝia endosimbionta Chlorella; aliaj pli provizoraj.
Multaj protistoj enhavas kaptitajn kloroplastojn, kloroplast-mitokondriajn kompleksojn, kaj eĉ okulilojn el algoj. La ksenosomoj estas bakteriaj endosimbiotoj troveblaj en ciliuloj, foje kun metanogena rolo ene de neaerobaj ciliuloj.[26]
Vidu ankaŭ
[redakti | redakti fonton]Notoj
[redakti | redakti fonton]- ↑ Cavalier-Smith, T. (2002). The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa (el Retarkivo 20150918231148). Int. J. Syst. Evol. Microbiol. 52, 297–354
- ↑ Cavalier-Smith, T. (2004). Chromalveolate diversity and cell megaevolution: interplay of membranes, genomes and cytoskeleton. En: Hirt, R.P., Horner, D.S. (Eds.). Organelles, Genomes and Eukaryote Phylogeny. CRC Press, Londres, pp. 75–108.
- ↑ Cavalier-Smith, T. (2007). Evolution and relationships of algae: major branches of the tree of life. En: Brodie, J., Lewis, J. (Eds.), Unravelling the Algae. CRC Press, Boca Raton, pp. 21–55.
- ↑ Cavalier-Smith, T. (2003). Protist phylogeny and the high-level classification of Protozoa. Eur. J. Protistol. 39, 338–348
- ↑ Cavalier-Smith, T. (2013) Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European Journal of Protistology 49 (2013) 115–178.
- ↑ Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., Cavalier-Smith, T., Guiry, M.D. kaj Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms.
- ↑ 7,0 7,1 7,2 Simpson, Alastair G. B.; Slamovits, Claudio H.; Archibald, John M. (2017). "Protist Diversity and Eukaryote Phylogeny". En Archibald, John M.; Simpson, Alastair G.B.; Slamovits, Claudio H. (eld.). Handbook of the Protists. Vol. 1 (dua eldono). Springer. pp. 1–22. doi:10.1007/978-3-319-28149-0_45. ISBN 978-3-319-28147-6.
- ↑ Simpson, Alastair G. B.; Slamovits, Claudio H.; Archibald, John M. (2017). "Protist Diversity and Eukaryote Phylogeny". En Archibald, John M.; Simpson, Alastair G.B.; Slamovits, Claudio H. (eld.). Handbook of the Protists. Vol. 1 (dua eldono). Springer. pp. 1–22. doi:10.1007/978-3-319-28149-0_45. ISBN 978-3-319-28147-6.
- ↑ Brown MW, Kolisko M, Silberman JD, Roger AJ (2012). "Aggregative Multicellularity Evolved Independently in the Eukaryotic Supergroup Rhizaria". Current Biology. 22 (12): 1123–1127. Bibcode:2012CBio...22.1123B. doi:10.1016/j.cub.2012.04.021. PMID 22608512. S2CID 17510471.
- ↑ Pawlowski, Jan (2008). "The twilight of Sarcodina: a molecular perspective on the polyphyletic origin of amoeboid protists" (PDF). Protistology. 5 (4): 281–302.
- ↑ Pawlowski, Jan; Burki, Fabien (2009). "Untangling the Phylogeny of Amoeboid Protists". The Journal of Eukaryotic Microbiology. 56 (1): 16–25. doi:10.1111/j.1550-7408.2008.00379.x. PMID 19335771.
- ↑ (2008) “Giant deep-sea protist produces bilaterian-like traces”, Current Biology 18 (23), p. 1849–1854. doi:10.1016/j.cub.2008.10.028. Bibkodo:2008CBio...18.1849M.
- ↑ (2019) “Giant protists (xenophyophores) function as fish nurseries”, Ecology 101 (4), p. e02933. doi:10.1002/ecy.2933.
- ↑ "Lynn Margulis replies". BioScience. 36 (5): 293–294. 1986. doi:10.1093/bioscience/36.5.293-a.
- ↑ Margulis, Lynn (1980). "Undulipodia, flagella and cilia". Biosystems. 12 (1–2): 105–108. Bibcode:1980BiSys..12..105M. doi:10.1016/0303-2647(80)90041-6. PMID 7378551.
- ↑ Andersen, R. A.; Barr, D. J. S.; Lynn, D. H.; Melkonian, M.; Moestrup, Ø.; Sleigh, M. A. (1991). "Terminology and nomenclature of the cytoskeletal elements associated with the flagellar/ciliary apparatus in protists". Protoplasma. 164 (1–3): 1–8. doi:10.1007/BF01320809.
- ↑ Thibaut Brunet; Marvin Albert; William Roman; Maxwell C Coyle; Danielle C Spitzer; Nicole King (la 15-a de januaro 2021), "A flagellate-to-amoeboid switch in the closest living relatives of animals" (in en), eLife 10, doi:10.7554/ELIFE.61037, ISSN 2050-084X, PMC 7895527, PMID 33448265, Vikidatumoj Q105870433
- ↑ Scamardella JM (1999). "Not plants or animals: A brief history of the origin of Kingdoms Protozoa, Protista, and Protoctista". International Microbiology. 2 (4): 207–221. PMID 10943416.
- ↑ (2021) “Protist diversity: Novel groups enrich the algal tree of life”, Current Biology 31 (11), p. R714–R740. doi:10.1016/j.cub.2021.04.025. Bibkodo:2021CBio...31.R733E.
- ↑ (2018) “What has happened to the "aquatic phycomycetes" (sensu Sparrow)? Part II: Shared properties of zoosporic true fungi and fungus-like microorganisms”, Fungal Biology Reviews 32 (2), p. 52–61. doi:10.1016/j.fbr.2017.09.003. Bibkodo:2018FunBR..32...52G.
- ↑ Neuhauser, Sigrid. (2012) “An introduction to fungus-like microorganisms”, Marine fungi, Marine and Freshwater Botany. De Gruyter, p. 137–152. doi:10.1515/9783110264067.137. ISBN 9783110264067.
- ↑ (1980) “A newly revised classification of the Protozoa”, Journal of Protozoology 27 (1), p. 37–58. doi:10.1111/j.1550-7408.1980.tb04228.x.
- ↑ (1993) “Kingdom Protozoa and its 18 phyla”, Microbiological Reviews 57 (4), p. 953–994. doi:10.1128/mr.57.4.953-994.1993.
- ↑ Facts about malaria (9a de Junio, 2017).
- ↑ (2018) “Evolutionary cell biology of proteins from protists to humans and plants”, J. Eukaryot. Microbiol. 65 (2), p. 255–289. doi:10.1111/jeu.12449. 206055044.
- ↑ 26,0 26,1 26,2 26,3 26,4 Levandowsky, Michael (2012). "Physiological Adaptations of Protists". En Sperelakis, Nicholas (eld.). Cell Physiology Sourcebook: Essentials of Membrane Biophysics (kvara eldono). Amsterdam; Boston: Elsevier/AP. pp. 874–890. ISBN 978-0-12-387738-3.
- ↑ Burki, Fabien; Sandin, Miguel M.; Jamy, Mahwash (2021). "Diversity and ecology of protists revealed by metabarcoding". Current Biology. 31 (19): R1267 – R1280. Bibcode:2021CBio...31R1267B. doi:10.1016/j.cub.2021.07.066. PMID 34637739. S2CID 238588753.
- ↑ 28,0 28,1 28,2 28,3 28,4 28,5 28,6 28,7 Adl, Sina M.; Bass, David; Lane, Christopher E.; Lukeš, Julius; Schoch, Conrad L.; Smirnov, Alexey; et al. (2019). "Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes". Journal of Eukaryotic Microbiology. 66 (1): 4–119. doi:10.1111/JEU.12691. PMC 6492006. PMID 30257078.
- ↑ 29,0 29,1 29,2 29,3 29,4 29,5 29,6 (1a de Julio 2024) “Feeding Mechanisms of Pathogenic Protozoa with a Focus on Endocytosis and the Digestive Vacuole”, Parasitologia 4 (3), p. 222–237. doi:10.3390/parasitologia4030019.
- ↑ 30,0 30,1 30,2 Hickman, Cleveland P. Jr.. (2017) “Unicellular Eukaryotes: Protozoan Groups”, Integrated Principles of Zoology, 7‑a eldono, New York: McGraw Hill, p. 216–245. ISBN 9781259562310. p. 217.
- ↑ (1946) “Nomenclature of nutritional types of microorganisms.”, Cold Spring Harbor Symposia on Quantitative Biology 11, p. 302–3.
- ↑ (November 1995) “Macropinocytosis”, Trends in Cell Biology 5 (11), p. 424–428. doi:10.1016/S0962-8924(00)89101-1.
- ↑ 33,0 33,1 (10 September 2013) “Horizontal gene transfer in osmotrophs: playing with public goods”, Nature Reviews Microbiology 11 (10), p. 720–727. doi:10.1038/nrmicro3108.
- ↑ (13a de Junio 2018) “Farming, slaving and enslavement: histories of endosymbioses during kinetoplastid evolution”, Parasitology 145 (10), p. 1311–1323. doi:10.1017/S0031182018000781.
- ↑ (2022) “Protozoan phagotrophy from predators to parasites: An overview of the enigmatic cytostome-cytopharynx complex of Trypanosoma cruzi”, Journal of Eukaryotic Microbiology 69 (6), p. e12896. doi:10.1111/jeu.12896.
- ↑ 36,0 36,1 36,2 Ruppert, Edward E.. (2004) “Protozoa”, Invertebrate Zoology: A Functional Evolutionary Approach, 7‑a eldono, Thomson Brooks/Cole, p. 22–57. ISBN 0-03-025982-7.
- ↑ 37,0 37,1 37,2 37,3 Esteban, Genoveva F.. (2020) “Feeding”, Ecology of Protozoa: The Biology of Free-living Phagotrophic Protists, 2‑a eldono, Cham: Springer Nature Switzerland AG, p. 33–54. doi:10.1007/978-3-030-59979-9_4. ISBN 978-3-030-59979-9.
- ↑ Geisen, Stefan; Koller, Robert; Hünninghaus, Maike; Dumack, Kenneth; Urich, Tim; Bonkowski, Michael (2016). "The soil food web revisited: Diverse and widespread mycophagous soil protists". Soil Biology and Biochemistry. 94: 10–18. Bibcode:2016SBiBi..94...10G. doi:10.1016/j.soilbio.2015.11.010.
- ↑ (16a de Junio 2015) “Pack-hunting protists attacking nematodes”, Environmental Microbiology 17 (11), p. 4538–4546. doi:10.1111/1462-2920.12949.
- ↑ (2011) “Alternative Nutritional Strategies in Protists: Symposium Introduction and a Review of Freshwater Protists that Combine Photosynthesis and Heterotrophy”, Journal of Eukaryotic Microbiology 58 (3), p. 181–184. doi:10.1111/j.1550-7408.2011.00543.x.
- ↑ (15 July 2019) “Mixotrophic protists and a new paradigm for marine ecology: where does plankton research go now?”, Journal of Plankton Research 41 (4), p. 375–391. doi:10.1093/plankt/fbz026.
- ↑ 42,0 42,1 Mitra, Aditee; Flynn, Kevin J.; Tillmann, Urban; Raven, John A.; Caron, David; Stoecker, Diane K.; Not, Fabrice; Hansen, Per J.; Hallegraeff, Gustaaf; Sanders, Robert; Wilken, Susanne; McManus, George; Johnson, Mathew; Pitta, Paraskevi; Våge, Selina; Berge, Terje; Calbet, Albert; Thingstad, Frede; Jeong, Hae Jin; Burkholder, JoAnn; Glibert, Patricia M.; Granéli, Edna; Lundgren, Veronica (2016). "Defining Planktonic Protist Functional Groups on Mechanisms for Energy and Nutrient Acquisition: Incorporation of Diverse Mixotrophic Strategies". Protist. 167 (2): 106–120. doi:10.1016/j.protis.2016.01.003. hdl:10261/131722. PMID 26927496.
- ↑ Faure, Emile; Not, Fabrice; Benoiston, Anne-Sophie; Labadie, Karine; Bittner, Lucie; Ayata, Sakina-Dorothée (Aprilo 2019). "Mixotrophic protists display contrasted biogeographies in the global ocean". The ISME Journal. 13 (4): 1072–1083. Bibcode:2019ISMEJ..13.1072F. doi:10.1038/s41396-018-0340-5. PMC 6461780. PMID 30643201.
- ↑ (21a de Marto 2023) “Euglenozoan kleptoplasty illuminates the early evolution of photoendosymbiosis”, Proceedings of the National Academy of Sciences of the United States of America 120 (12), p. e2220100120. doi:10.1073/pnas.2220100120. Bibkodo:2023PNAS..12020100K.
- ↑ (18a de Januaro 2022) “Trophic flexibility of marine diplonemids - switching from osmotrophy to bacterivory”, The ISME Journal 16 (5), p. 1409–1419. doi:10.1038/s41396-022-01192-0. Bibkodo:2022ISMEJ..16.1409P.
- ↑ 46,0 46,1 (February 1980) “Contractile vacuoles and associated structures: their organization and function”, Biological Reviews 55 (1), p. 1–46. doi:10.1111/j.1469-185x.1980.tb00686.x.
- ↑ (25a de Majo 2009) “Molecular phylogeny of ocelloid-bearing dinoflagellates (Warnowiaceae) as inferred from SSU and LSU rDNA sequences.”, BMC Evolutionary Biology 9 (1), p. 116. doi:10.1186/1471-2148-9-116. Bibkodo:2009BMCEE...9..116H.